

A good site for collecting y-larvae: Gongguan Fishing Harbor, Green Island, Taitung County. Photo by Dr. Grygier.適合採集 y 型幼生的地點:臺東縣綠島公館漁港(Dr. Grygier攝)
文、圖 Authors-
‧Mark J. Grygier, Distinguished Researcher, National Museum of Marine
Biology & Aquarium,Taiwan
國立海洋生物博物館特聘講座
‧Jørgen Olesen, Associate Professor, Natural History Museum of
Denmark
丹麥自然歷史博物館副教授
‧Niklas Dreyer, Genomics Specialist, Centre de recherche du CHU
Sainte-Justine, Université de Montréal, Canada
蒙特婁大學聖賈斯汀醫學中心基因組學專家
中文翻譯與審稿 Translation and Review-
‧Chinese Translation – Coral Hsieh (謝佳珊)
‧Chinese Review – Benny K.K. Chan (陳國勤), Research Fellow and Director, Biodiversity Research Center,
Academia Sinica, Taiwan
中央研究院生物多樣性研究中心研究員及主任
Many microscopic creatures discovered
during the early days of marine plankton research were eventually shown to be
larvae— infant stages in the life history— of familiar creatures like crabs and
sea urchins. By now, almost all the planktonic larval forms have been matched
to adult marine organisms, but one diverse group of crustacean larvae still
lacks any recognized adults: the y-larvae. Despite their mysterious nature,
y-larvae have recently been shown to be very abundant at certain places in East
Asia, including Taiwan, and therefore may be of ecological importance.
Their story starts in 1899, when the Danish
zoologist H. J. Hansen published a report on planktonic barnacle larvae from
the Atlantic Ocean. Barnacles have two kinds of larvae. The nauplius larva has
three pairs of limbs (first and second antennae and mandibles) and also a pair
of anterolateral horns that distinguish barnacle nauplii from any other kind of
crustacean nauplius. After molting several (usually six) times, the nauplius
becomes a cypris: a non-feeding, shell-enclosed creature with six pairs of
swimming legs and a pair of specialized antennae with which it attaches itself
to a rock or some other hard substrate. The attached cypris metamorphoses into
a filter-feeding barnacle, which, upon reaching maturity, eventually releases
more nauplius larvae into the plankton. Y-larvae also have nauplius and cypris
stages, but these differ from their barnacle counterparts.
What are y-larvae? In his 1899 report,
Hansen described and illustrated several kinds of nauplius that seemed related
in some way to barnacles but lacked anterolateral horns and had a complex
pattern of dorsal cuticular ridges. He called them "Nauplius y", and
such larvae are now commonly called Hansen’s y-larvae. In formal zoological
classification, they constitute the crustacean subclass Facetotecta; molecular
systematics has confirmed that this is a valid, monophyletic group that
represents the first branch of the phylogenetic tree leading to barnacles. Up
through the 1970s, just a few additional records of nauplius y were published
from the North Atlantic and the Mediterranean Sea, as well as the first
discoveries of planktonic cypris y larvae off Denmark and the Bahamas. Between
1984 and 1991 the Japanese carcinologist Tatsunori Itô described numerous forms
of both nauplius y and cypris y from Tanabe Bay, Honshu. He established the
genus Hansenocaris for them, showed that the nauplius molts at least five
times, and devised a nomenclature to describe the ridge-bounded “plates” or
“facets” of the naupliar cephalic shield.
.jpg)


The most common morphospecies of y-larva in Bisha Fishing Harbor, Keelung: Last - stage nauplius y (ventral and dorsal views of molted cuticle) and cypris y (lateral view). SEM photos by Dr. Dreyer.
基隆碧砂漁港最常見的Y幼體:末期無節幼生y(脫落外骨骼的腹面和背面視圖)及絲足幼生Y側面視圖)(掃描電子顯微鏡圖像由Dr. Dreyer拍攝)
How can we recognize a facetotectan
nauplius or cypris? Unlike barnacle nauplii, they have no anterolateral horns,
and unlike copepod nauplii they have no hair-like caudal setae, just three
posterior spines. They have a dome-like cephalic shield, usually ornamented
with a turtle-shell-like reticulation, and the ventral side of the head outside
the bases of the limbs is wide and flat. The distinct trunk region tapers
either sharply or gradually and the ventral side is often somewhat inflated.
Great variation is seen in the labrum (upper lip) and the caudal armament as
well as the overall body form; some y-nauplii resemble helmets, Chinese soup
spoons, airships, discs, or tennis rackets, among other shapes. Y-nauplii with
complexly armed second antennae and mandibles and a plate-like labrum evidently
feed in the plankton (planktotrophic nauplii), but their preferred food is
still unknown. Other y-nauplii that have
little or no armament besides swimming setae on their limbs can undergo a
complete sequence of molts in the laboratory without feeding (lecithotrophic
nauplii, see photo above).
A cypris y has a cephalic shield or
carapace that is shaped like an upside-down boat hull but does not enclose the
whole body like that of a barnacle cypris. The first antennae and labrum
usually bear hooks. The six pairs of paddle-like thoracic swimming legs are
followed by a two- or (much more usually) four-segmented abdomen with a bulky
terminal segment. The shield and various body surfaces are often distinctly
ridged. Most kinds of cypris y are swift swimmers. The life history stage
following the cypris, the limb-less, slug-like, non-feeding ypsigon larva, is
known mostly through artificial exposure of cypris y larvae to crustacean
molting hormone. The ypsigon may be the host-invasive stage if adult y-larvae
turn out to be parasitic.
Most recent work on y-larvae has been done
by two overlapping research teams with members from Taiwan, Denmark, Russia,
Spain, and Japan, including the present authors. For more information about
Facetotecta in general, please see their recent review paper.[1] Our team’s work
on East Asian y-larvae so far has mainly concerned non-feeding nauplius larvae
from Sesoko Island in Okinawa and from Taiwan. Our study material includes
about 14,000 specimens collected since 2017, as well as older Okinawan material
going back to 1996. Among them, the number of ‘morphospecies’ recognizable by
eye is considerably less than the number distinguished by the nucleotide
sequences of various genes. A paper in 20232 tentatively recognized 34 naupliar
morphospecies (8 planktotrophic and 26 lecithotrophic) that appeared to
represent at least 88 genetic species. Of the 34, 22 were only from Okinawa, [2] only from Taiwan, and 9 from both places. In 2024, two of the present authors
added to the Okinawan total by diagnosing and illustrating a total of 34
lecithotrophic naupliar morphospecies from Sesoko Island, while also noting the
presence of at least 7 others there plus the above-mentioned planktotrophic
forms.[3]
Formal taxonomy of these collections has
barely begun, with only three Okinawan species having been formally described
with Latin names. The intention, however, is to base new taxonomy on sets of
laboratory-reared last naupliar stages and the cypris larvae that molted from
them, so that 1) equivalent stages can serve as the basis for comparison among
species and 2) there can be no mistake about the conspecificity of both stages
for any particular species. We intend to gather as much information as possible
from different stages of single specimens using an assortment of techniques
like videography, molecular analysis, scanning electron microscopy, and light
microscopy augmented by image-stacking and autofluorescence-based confocal
laser scanning, as shown in our YouTube videos.[4]
Single-larva DNA barcoding using both
mitochondrial and nuclear genes has already shown that non-feeding forms of
nauplius y have arisen several times in phylogeny.2 It also indicates the
inadequacy of the current taxonomy, in which Hansenocaris, currently with 17
named species, is still the only recognized genus of y-larvae. A para-taxonomy
of alphabetically-labeled ‘types’ is being used as a stop-gap for all the East
Asian forms while they await formal description. The sheer number of sympatric
morphospecies of y-larvae confirmed at different sites (almost 50 at Sesoko
Island alone!) poses a severe problem, not only taxonomic but also ecological:
where are the adults hiding, and what should we be looking for? This problem
remains even if we assume the adults are parasitic. What creatures are likely
to host such a diversity of undiscovered parasites?
Where and how can a person catch y-larvae
in Taiwan? The easiest way is by tossing an ordinary 100-µm mesh plankton net
multiple times from a pier or wharf, by towing the net by hand alongside the
wharf, or by pouring buckets of seawater through the net, and then sorting the
resulting sample under a back-lit dissecting microscope. The numbers
encountered per sample will range from zero up to 50 or 100 or more. Y-larvae
can be recovered from both living and preserved samples, but when possible,
some living specimens of lecithotrophic nauplius larvae should be reared to the
cypris stage, a process that may take a few days to two weeks depending on
species. The last-stage nauplius y is recognizable by the compound eyes of the
cypris y developing within it.
The main sampling site for our published
Taiwanese material has been Green Island (Gongguan Fishing Harbor), but they
are also consistently present in low numbers in Bisha Fishing Harbor, Keelung.
A plankton survey conducted mainly in September 2019 and March to November 2020
at 52 other fishing harbors, mostly in northern and southern Taiwan and in
Penghu, turned up other good sites. Y-larvae were most abundant at Wanghaiziang
Fishing Harbor, Keelung; Fenniaolin and Wushi Fishing Harbors, Yilan; southern
Xiaoliuqiu Island, Pingtung; and several fishing harbors around Xiyu Island,
Penghu. Much of the coastline remains unexplored, however. Eventually, it would
be nice to produce a guidebook showing the complete inventory of Taiwan’s
y-larvae, similar to our photographic catalogue of Okinawan lecithotrophic
y-naupliar morphospecies.[3]
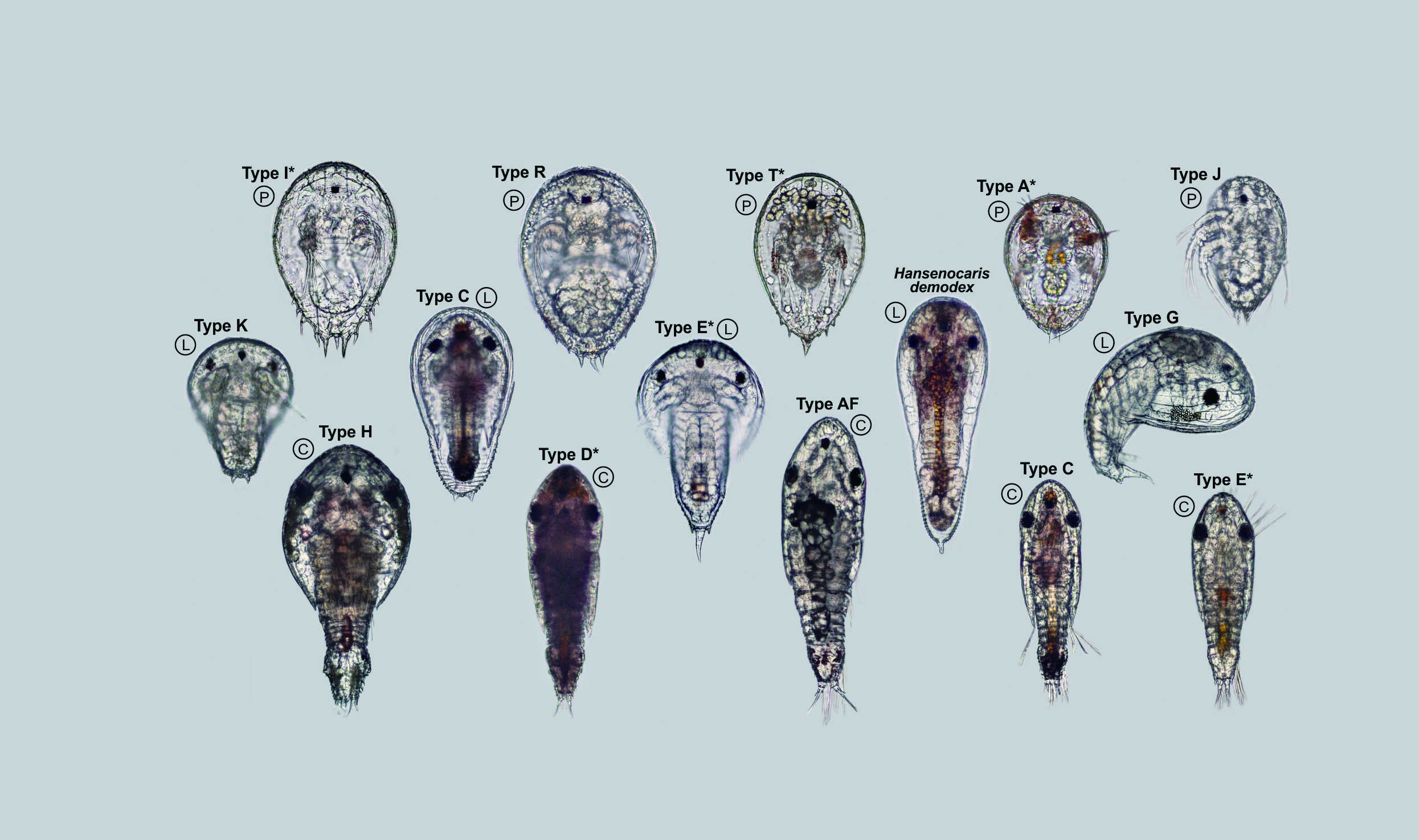
Several letter-designated “Types” (morphospecies) of planktotrophic Ⓟ and last-stage lecithotrophic Ⓛ nauplius y as well as cypris y Ⓒ from Sesoko Island, Okinawa, Japan. Photos by Jørgen Olesen, all to same scale.
來自日本沖繩瀨底島的形態物種,包括浮游性 Ⓟ 型和末期卵黃性 Ⓛ 型幼蟲Y,以及圓形幼蟲Y Ⓒ,照片由Dr. Olesen 拍攝,所有照片比例相同
許多在海洋浮游生物研究早期被發現的微生物,後來都被證實是某些大家耳熟能詳物種的幼體,例如螃蟹或是海膽。隨著研究發展至今,幾乎所有浮游幼體都能找到特定的海洋物種成體。但現今還有一大群無法被辨識的甲殼類幼體,也就是本篇要討論的Y幼體。儘管生命週期依然是個謎,牠們最近卻大量出現在包含臺灣海域的東亞地區,也因此被認為具有潛在的生態重要性。
Y幼體的故事始於1899年,當時丹麥動物學家H. J. Hansen發表了一份關於大西洋海域藤壺幼體的報告。通常藤壺有兩個幼體階段:一開始的無節幼體階段(nauplius)有3對附肢(第一和第二對觸角及下顎),以及一對前側角,這些特徵使得藤壺的無節幼體有別於其他甲殼類的無節幼體。經過6次左右的蛻皮後,無節幼體會變成腺介幼體:一種不進食且被兩片幾丁質的殼包覆住的階段。腺介幼體具有6對泳足和一對特化的觸角,腺介幼體便是利用這對觸角將自己附著在岩石或其他堅硬的基質上。附著在岩石上的幼體會蛻變成為濾食性的藤壺成體,在成熟後釋放更多的無節幼體於大量的浮游生物中。Y幼體也有無節幼體和腺介幼體這兩個階段,但牠們並沒有前側角,還是與藤壺的幼體有所區隔。
那麼Y幼體究竟是什麼呢? Hansen在 1899年的報告中描述並繪製了幾種似乎與藤壺有某種關聯的無節幼體,這些幼體缺乏前側觸角,而且在背部具有複雜的角質隆起圖紋。他無法辨認此類幼體,故將牠們稱為「 Y無節幼體」。這些幼體現在則通常被稱為 Hansen 氏 Y幼體。在正式的動物分類中,牠們屬於甲殼類亞綱「帶甲下綱」Facetotecta;分子系統分類學上已經證實這是一個有效的單系群,代表著導向藤壺這類生物的系統樹上第一個分支。直到1970年代,只有在北大西洋和地中海地區發表少數 Y 無節幼體的其他記錄,並首次在丹麥和巴哈馬發現了浮游性Y 腺介幼體。從1984年到1991年,日本甲殼類學家伊藤立則描述了位於本州田邊灣裡的多種 Y 無節幼體和 Y腺介幼體。他為這些幼體建立了 Hansenocaris 屬,證明這個屬的無節幼體至少蛻皮5次,並設計一套命名法以描述這些無節幼體頭盾上隆起的板狀或面狀範圍。
進一步來說,如何辨認帶甲下綱的無節幼體或腺介幼體?與藤壺的無節幼體不同的是,牠們沒有前側角。且與橈足類的無節幼體不同之處在於,牠們沒有髮狀的尾部剛毛,只有3根位於後端的脊柱。帶甲下綱的無節幼體有一個圓頂狀的頭部甲殼,通常帶有像龜殼一樣的網狀紋路,且在頭的底部、肢體基部外側顯得寬而平。軀幹部位呈現有特色的尖銳或平緩的錐狀,也時常可以發現到些微膨脹的腹面。上唇板和尾部武器般的甲板,以及整體體型也存在著很大的變異;一些 Y 幼體看起來像頭盔、中式湯匙、飛船、圓盤或網球拍等形狀。牠們具有複雜剛毛的第二對觸角和大顎,以及扁平唇板的Y無節幼體顯然會取食浮游生物(浮游營養型無節幼體),但科學家仍未發現牠們偏好取食何種浮游生物。另一種除了肢體上的游泳剛毛外,幾乎沒有複雜剛毛的Y無節幼體,則可以在實驗室中以不進食的狀態下經歷完整的蛻皮過程(卵黃營養型無節幼體)。
Y 腺介幼體頭部的背甲形狀像一艘顛倒的船,但不像藤壺的腺介幼體那樣包覆全身。牠的第一對觸角和上唇板通常帶有鉤狀結構。有著6對槳狀泳足的胸部,後面接著由2節或更常見4節組成的腹部,而最後一節則顯得特別厚實。牠們的背甲和多樣的身體表面通常有明顯的脊狀結構,而且大多數 Y 腺介幼體游速很快。腺介幼體之後的生命階段是 ypsigon 幼體,在這個時期牠們沒有明顯的肢體,長得像蛞蝓一樣,也不進食。這個階段主要是由於人為因素,Y腺介幼體暴露在甲殼類動物的脫殼激素中而形成。如果成年後是寄生性的Y 幼體,那麼 ypsigon 幼體則可能是侵入宿主的階段。

最近關於 Y 幼體的研究主要還是由成員來自臺灣、丹麥、俄羅斯、西班牙和日本的兩個研究團隊進行,其中包括本文的作者。關於「帶甲下綱」的更多資訊,請參閱上述團隊最近的綜述論文[1]。本團隊對東亞海域的 Y 幼體研究目前主要集中在沖繩瀨底島和臺灣海域中不進食狀態的無節幼體。我們的研究材料除了從2017年以來蒐集到約14,000件標本,還有可以追溯到1996年自沖繩取得較舊的材料。其中,用肉眼就能辨別的「形態種」數量明顯少於藉由不同基因的核苷酸序列區分出的種類。2023年的一篇論文初步確認了34種無節幼體形態種(8種浮游型和26種卵黃營養型)[2],這些形態種似乎代表了至少88個基因物種。在這34種形態種中,22種僅在沖繩瀨底島發現,2種來自臺灣,9種則在兩地都有被發現。2024年,本文其中的兩位作者增補了沖繩的總數,除了確認並繪製瀨底島的34種卵黃營養型無節幼體形態種之外,並指出至少還有7種其他形態種以及上述的浮游性形態種存在。
關於這些物種的正式分類學仍處於初始階段,目前只有3種沖繩物種已經以拉丁學名正式命名。這個計畫的目的是:以在實驗室飼養的最後無節幼體階段以及從這些幼體蛻變而來的腺介幼體為基礎,建立新的分類學。這可以使:1. 相同階段可作為物種之間比較的基礎,2. 對於任何特定物種的兩個幼體階段之間的同種性不會有所出入。我們希望盡可能利用各種技術,從單一標本上蒐集到不同發育階段最多的資訊,像是攝影、分子分析、掃描電子顯微鏡和光學顯微鏡,並輔以疊圖和自體螢光共焦雷射掃描,而這些技術也展示在我們的YouTube影片中[4]。

Author Mark J. Grygier sampling y-larvae with a plankton net at Haizikou Fishing Harbor, Xiaoliuqiu Island, Pingtung County. Photo by Chang Yu Shan.
作者 Mark J. Grygier 在屏東縣小琉球島海子口漁港使用浮游生物網採集 Y-幼體,張瑀珊攝
透過粒線體和細胞核內基因進行單一幼蟲的DNA解碼過程已經顯示,無攝食型態的Y幼體在親緣關係中已多次出現。而這也顯示了當前分類系統的不足,現有17個命名物種的Hansenocaris仍是唯一被承認的Y幼體屬。目前,對於所有東亞型態的Y幼體,皆使用以字母標記的「類型」作為臨時分類系統,尚待進一步正式的描述。不同地點確認的共域Y幼體形態種數量驚人(光是在瀨底島就有近50種!),這不僅對分類學構成嚴峻挑戰,對生態學也是如此:牠們的成年個體究竟躲在哪裡?我們應該要找的究竟是什麼?即便假設成體是寄生生物,這個問題依然存在:什麼生物會是這些尚未被發現的多樣寄生物的宿主?
在臺灣,可以在哪裡以及如何捕捉Y幼體呢?最簡單的方法是從碼頭或港口多次拋出普通的100微米網目浮游生物網,或沿著碼頭手動拖曳濾網,又或者將海水一桶一桶倒入網中過濾,接著在背光解剖顯微鏡下篩選樣本。每次取樣過程中,可蒐集到的Y幼體數量從0到50或100以上。Y幼體可以從活樣本或保存樣本中取得,但在可能的情況下,應該將部分具有卵黃營養型無節幼體飼養到腺介幼體階段,這個過程可能依照物種的不同,需要花上幾天到兩週不等的時間。最後階段的Y無節幼體可以藉由其中正在發育的Y腺介幼體的複眼加以識別。
關於我們已經發表的臺灣樣本,主要採樣地點是綠島的公館漁港,但在基隆的碧沙漁港也能發現穩定的少量紀錄。2019年9月及2020年3月至11月期間,在主要分布於臺灣北部、南部和澎湖的52個漁港中所進行的浮游生物調查,也發現其他良好的採樣地點。Y幼體在基隆望海巷漁港、宜蘭粉鳥林和烏石漁港、屏東小琉球南部以及澎湖西嶼島周圍的幾個漁港最為豐富。但其實大部分臺灣周遭的海岸線仍未被好好探索。最後,希望未來能製作一本關於臺灣海域Y幼體的完整型態資料的指南,類似我們針對沖繩的卵黃營養型Y無節幼體形態種所做的影像紀錄[3]。

Last-stage nauplius y (living) and cypris y (preserved) of same laboratory-reared specimen of Type C from Okinawa (not to same scale)
來自沖繩同一實驗室培養樣本的末期無節幼體幼體 Y(活體)和C型態的圓形幼蟲 Y(保存標本)(比例不一)
參考資料(REFERENCES):
1 Dreyer, N., Olesen, J., Grygier, M. J.,
Eibye-Jacobsen, D., Høeg, J. T., Kerbl, A., Fujita, Y., Kolbasov, G. A.,
Savchenko, A. S., Worsaae, K., Palero, F. & Chan, B. K. K. (2023). The
biology and lifecycle of enigmatic crustacean y-larvae: a review. Oceanography
and Marine Biology: An Annual Review, 61:81-126, https://doi.org/10.1201/9781003363873-4.
2
Dreyer, N., Palero, F., Grygier, M.J., Chan, B.K.K. & Olesen, J.
2023. Single-specimen systematics resolves the phylogeny and diversity
conundrum of enigmatic crustacean y-larvae. Molecular Phylogenetics and Evolution,
184:107780, https://doi.org/10.1016/j.ympev.2023.107780.
3
Olesen, J. & Grygier, M.J. 2024. Taxonomic diversity of marine
planktonic ‘y-larvae’ (Crustacean: Facetotecta) from a coral reef hotspot
locality (Japan, Okinawa), with a key to y-nauplii. European Journal of
Taxonomy. 929:1-90, https://doi.org/10.5852/ejt.2024.929.2479.
4 https://youtu.be/seo-63AK10E, https://youtu.be/SCskuhPTCXo,
https://youtu.be/er0mYLswV-c, https://youtu.be/u0megFZ6OTc
沒有留言:
張貼留言